溶瘤病毒治疗,尽管具有针对肿瘤细胞的选择性复制和杀伤能力,但其疗效受到一些限制。这些局限性包括系统给药时,病毒在肿瘤组织中的吸收不足,以及病毒可能导致免疫检查点蛋白过度表达,从而抑制免疫系统的反应。肿瘤微环境对治疗效果的影响也是一个关键因素。在微环境中,免疫检查点蛋白如PD-L1的过表达可以降低免疫反应,进而减少治疗效果。
为了应对这些挑战,基因编辑技术,特别是CRISPR/Cas9,被提出用于破坏肿瘤细胞中的免疫检查点基因。这种策略有望逆转肿瘤微环境的免疫抑制状态,从而提高治疗效果。
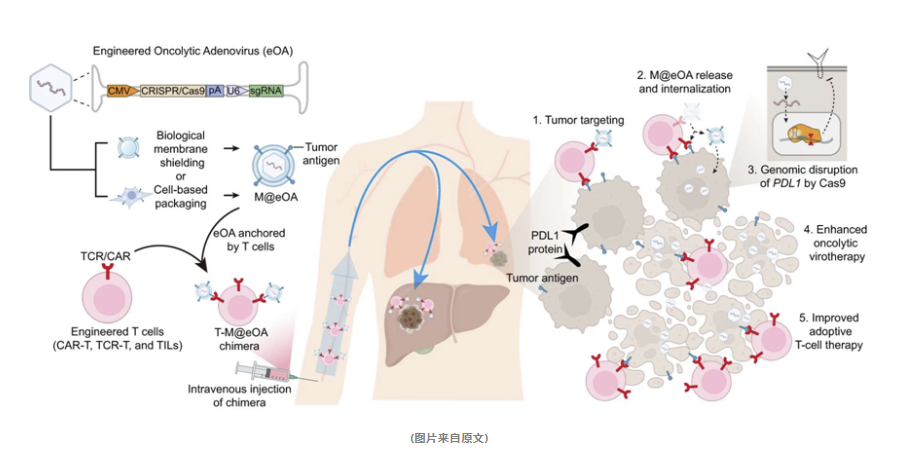
然而,肿瘤靶向给药仍然面临挑战,例如需要开发更有效的载体系统,以确保药物能够准确地送达肿瘤部位。一种有前景的策略是将溶瘤病毒负载在具有肿瘤趋向性的免疫细胞表面,以实现病毒和细胞的同步给药,这有望提高治疗效果。此外,开发溶瘤病毒基因编辑治疗策略,可以克服当前溶瘤病毒治疗存在的局限性,为肿瘤治疗提供新的解决方案。为此,浙江大学药学院平渊团队开发了一种名为ONCOTECH的新型治疗方法,该方法作为一种结合病毒治疗和细胞治疗的技术,展现了良好的临床转化潜力。该研究于2024年2月9日在线发表于Nature Biotechnology期刊(IF:46.9),论文标题为“An oncolytic virus–T cell chimera for cancer immunotherapy”。

在这项创新性的研究中,研究团队采取了一种独特的方法,将肿瘤溶解病毒(具体来说是肿瘤溶解腺病毒,简称为OAs)与T细胞相结合,以期提升对癌症的治疗效率。具体来说,该方法(ONCOTECH)利用了一种携带Cas9基因和靶向PDL1 sgRNA的eOA病毒,通过精准的基因编辑技术,永久性地下调肿瘤细胞中PDL1的表达,从而增强溶瘤病毒的疗效。
此外,将eOA病毒装载到T细胞表面,形成了一个复合体,实现了eOA病毒和T细胞的同步靶向给药,这不仅增强了T细胞的治疗效能,还改善了肿瘤免疫抑制微环境。ONCOTECH能够降低肿瘤和免疫细胞上PDL1的表达,增加效应T细胞的浸润,同时减少调节性T细胞的数量,这一系列作用共同提升了免疫治疗的效果。
更进一步,eOA病毒的感染能够诱导肿瘤细胞发生免疫原性死亡,释放危险信号分子,激活免疫系统。同时,ONCOTECH还能够促进树突细胞提呈肿瘤抗原,激活肿瘤浸润T细胞,从而增强T细胞的治疗效果。
01
eOA 释放的 Cas9 如何有效破坏肿瘤细胞 PDL1 基因?
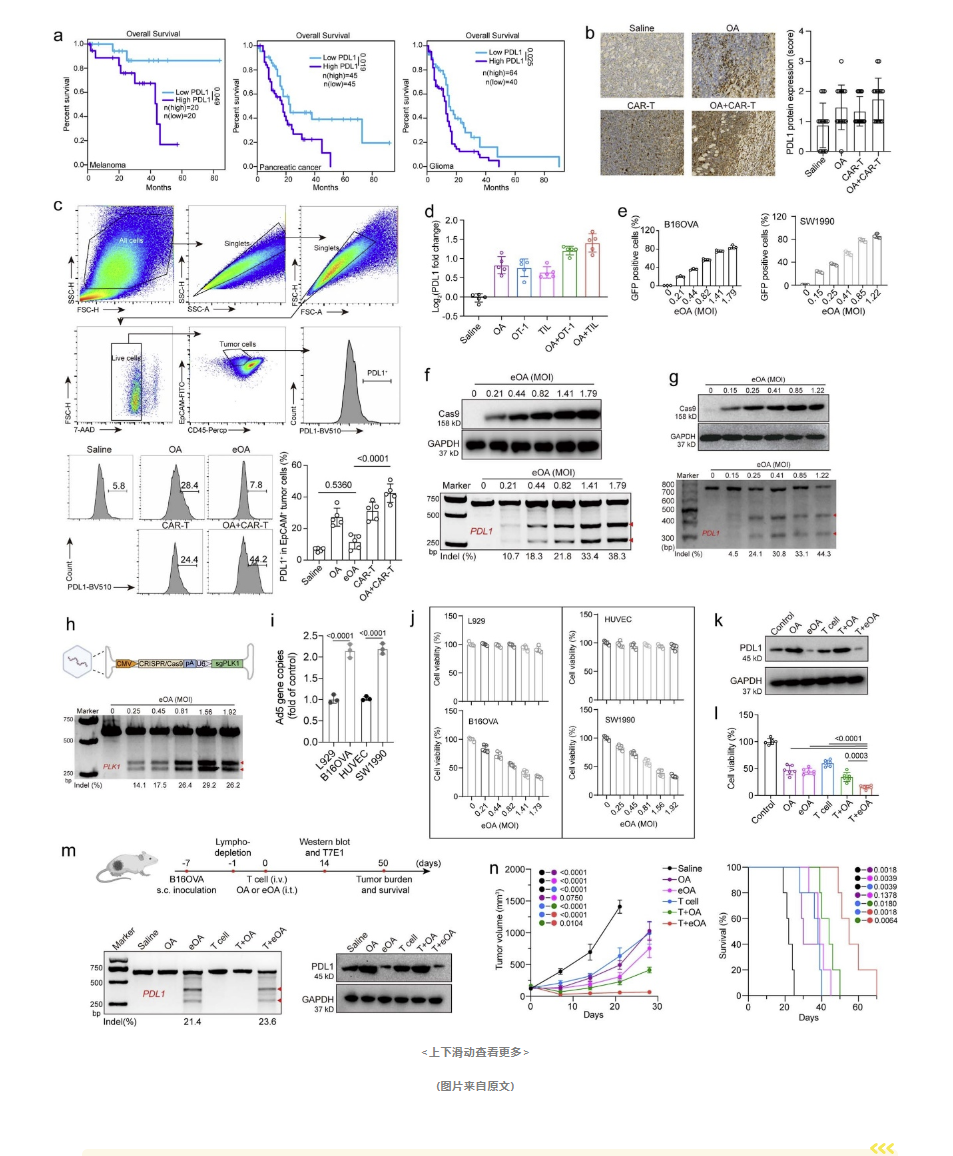
在该研究中,研究人员设计了一种溶瘤腺病毒(OA),命名为eOA,该病毒携带了Cas9基因编辑系统以及针对PDL1基因的特定sgRNA。这一创新的基因编辑策略旨在直接破坏肿瘤细胞中的PDL1基因,从而提升免疫系统对癌细胞的攻击能力。
研究人员首先使用eOA感染肿瘤细胞,并观察到Cas9蛋白及其伴随的sgRNA被成功转录到肿瘤细胞内。通过T7E1基因编辑检测,他们发现PDL1基因发生了预期的插入和缺失突变,这表明Cas9已经准确地编辑了目标基因。
进一步的Western blot检测显示,eOA感染后,肿瘤细胞中的PDL1蛋白表达显著下降。此外,通过T7E1和Western blot检测,研究人员还证实了eOA能够在体内环境中破坏肿瘤细胞的PDL1基因,并有效下调PDL1蛋白的表达。
02
如何巧妙地将病毒颗粒装载到载体 T 细胞膜上?
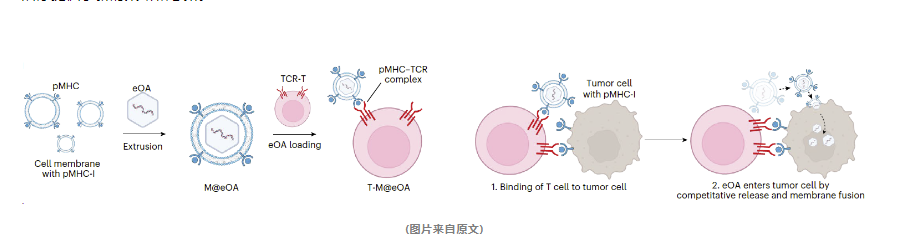
在该项研究中,研究人员采用了一种创新的技术手段,成功地将病毒颗粒有效装载到了T细胞的表面。这一过程涉及了精心的设计和多个步骤:
研究团队首先利用脂质挤出法,将工程化的腺病毒(eOA)封装到表达MHC-I-卵清蛋白复合物的B16OVA细胞膜中,从而形成了被称为M@eOA的复合体。
接下来,研究人员利用MHC-I-卵清蛋白复合物与OT-1 CD8+ T细胞上的TCR受体之间的特异性识别作用,实现了M@eOA复合体在OT-1 T细胞表面的锚定,进而形成了T-M@eOA复合体。
值得注意的是,M@eOA是通过脂质双分子层锚定在T细胞膜上,而非通过共价键连接,这样的方式确保了T细胞的功能不会受到影响。
当T-M@eOA复合体到达肿瘤细胞并识别出肿瘤特异性抗原时,M@eOA能够与T细胞竞争性地释放到肿瘤细胞表面,并成功进入肿瘤细胞内部。这种基于抗原受体相互作用的生物物理方法,不仅能够有效地将病毒颗粒装载到T细胞膜上,而且为肿瘤靶向的病毒治疗提供了新的策略和思路。
03
ONCOTECH如何增强体外抗肿瘤反应?
1
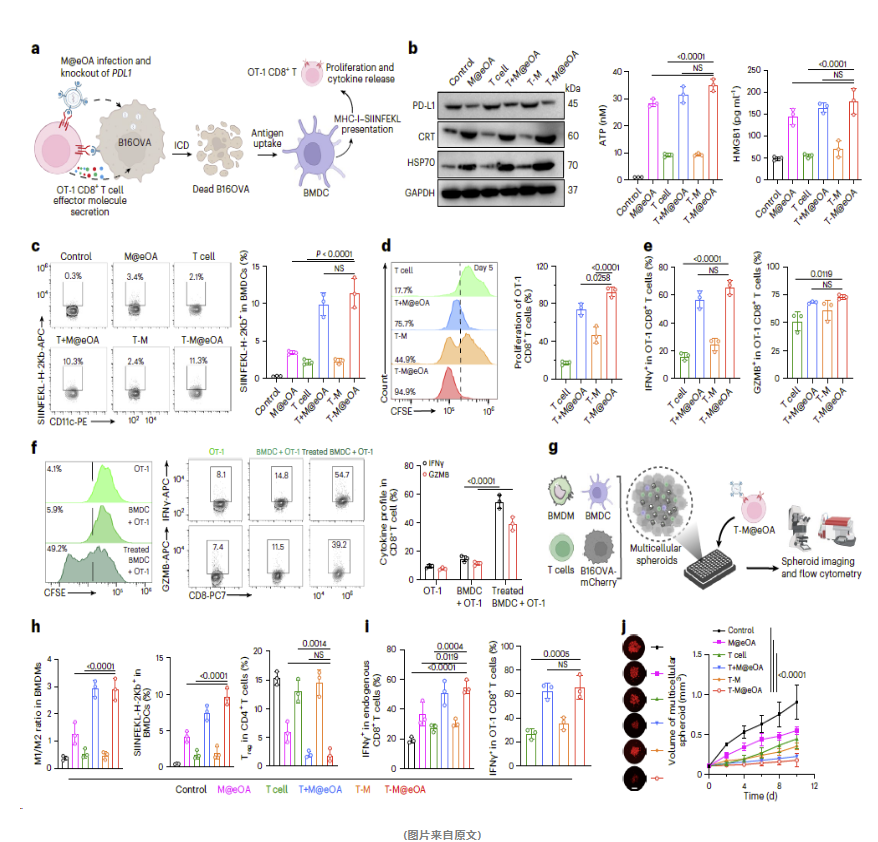
免疫细胞死亡(ICD)标记物的表达增加
当肿瘤细胞经过T-M@eOA处理后,免疫细胞死亡(ICD)的标记物,包括钙网蛋白(CRT)、热休克蛋白70(HSP70)、ATP和HMGB1的表达水平显著上升。这一现象表明T-M@eOA能够诱发强烈的ICD效应,从而激活免疫系统。
2
抗原提呈能力的增强
实验显示,经T-M@eOA处理的肿瘤细胞上清液可以增强骨髓来源的树突状细胞(BMDC)对MHC-I-卵清蛋白复合物的提呈能力。这意味着T-M@eOA能够提升抗原提呈细胞的功能,进一步增强免疫应答。
3
T细胞增殖和效应功能的增强
T-M@eOA能够显著促进T细胞的增殖,并提高其分泌干扰素γ和颗粒酶B等效应分子的能力,这些都是评估T细胞功能的重要指标。
4
肿瘤球生长的抑制
在三维肿瘤球模型中,T-M@eOA能够有效抑制肿瘤球的生长,这表明它能够重塑免疫抑制的肿瘤微环境,并增强T细胞的抗肿瘤功能。
5
基因集富集分析
通过对T-M@eOA处理后的肿瘤组织进行RNA测序和基因集富集分析,研究人员发现病毒蛋白与细胞因子及其受体相互作用的通路被激活,这进一步揭示了ONCOTECH作用的潜在分子机制。
综合以上实验结果,研究人员得出结论,ONCOTECH通过多种机制重塑了免疫抑制的肿瘤微环境,并增强了T细胞的抗肿瘤功能,从而提高了抗肿瘤免疫反应的效果。
04
ONCOTECH如何增强体外抗肿瘤反应?
在该研究中,关键的实验结果表明ONCOTECH疗法通过多种机制改善免疫抑制性TME,激活免疫应答,并增强抗肿瘤免疫反应:
1
免疫抑制性TME改善
经过T-M@eOA治疗的小鼠肿瘤组织中,PD-L1的表达显著降低。这一变化表明ONCOTECH疗法有助于缓解肿瘤微环境的免疫抑制状态。
2
ICD效应增强
治疗后的肿瘤细胞显示出更高的钙网蛋白(CRT)、热休克蛋白70(HSP70)、ATP和HMGB1的表达水平,这些都是免疫细胞死亡(ICD)效应的关键标记物。增强的ICD效应有助于激活免疫系统对肿瘤细胞的攻击。
3
抗原提呈能力提升
T-M@eOA治疗不仅增加了成熟树突细胞(DC细胞)的比例,还增强了DC细胞对SIINFEKL抗原的提呈能力,这对于启动和维持有效的免疫反应至关重要。
4
免疫细胞数量增加
治疗后,小鼠肿瘤组织中的M1样巨噬细胞和内源性CD8+ T细胞数量显著增加,而调节性T细胞(Treg细胞)的数量减少。这种变化有助于创建一个更有利于免疫介导的肿瘤杀伤的环境。
5
免疫应答激活
通过对T-M@eOA处理后的肿瘤组织进行基因集富集分析,研究人员发现多种与免疫相关的通路被激活,包括病毒蛋白与细胞因子受体相互作用等关键通路。
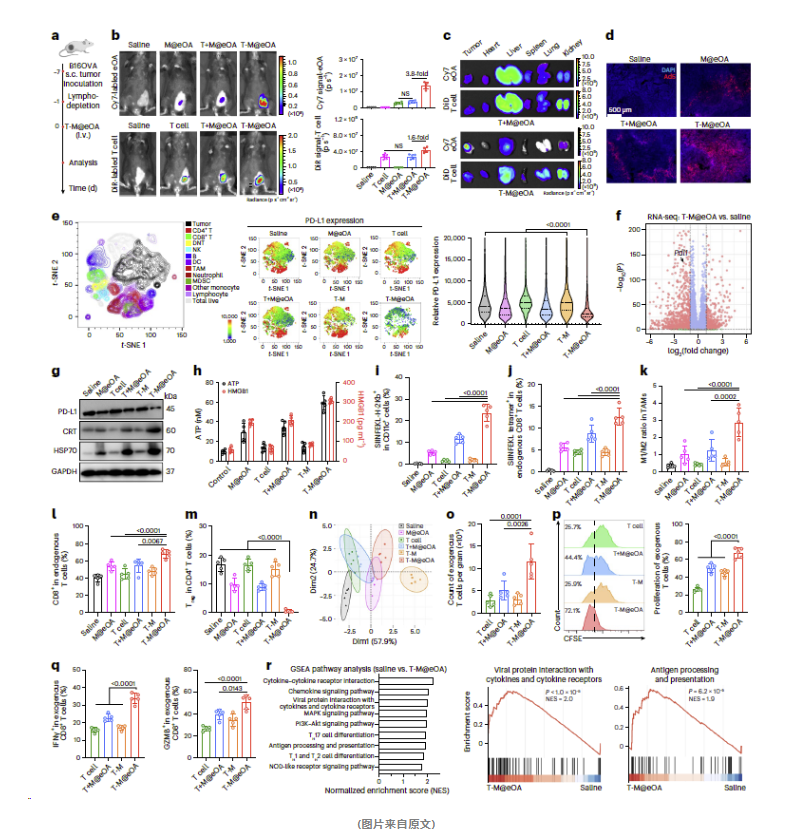
05
ONCOTECH治疗小鼠实体瘤效果如何?
在一系列深入的实验研究中,研究人员发现了ONCOTECH疗法在治疗实体瘤方面的显著效果。以下是关键的实验结果:
1
小鼠黑色素瘤模型
在小鼠皮下B16OVA肿瘤模型中,T-M@eOA治疗显著抑制了肿瘤的生长,并且明显延长了小鼠的生存期。
2
转移性肿瘤模型
针对B16OVA转移性肿瘤模型,T-M@eOA治疗不仅显著抑制了肿瘤的生长,还明显减少了肺部转移结节的数量,并提高了小鼠的生存期。
3
自发转移肿瘤模型
在4T1-OVA自发转移肿瘤模型中,T-M@eOA治疗同样有效地抑制了肿瘤的生长,减少了肺部转移结节的数量,并提高了生存期。
4
人源化PDX模型
在更为临床相关的人源化患者衍生肿瘤(PDX)模型中,T-M@eOA治疗显著抑制了肿瘤的生长,并且延长了生存期。
5
患者来源肿瘤细胞
实验还表明,eOA能够成功地装载到患者来源的肿瘤浸润淋巴细胞(TIL细胞)上,并显著杀死肿瘤细胞。
综合这些实验结果,研究团队得出结论:ONCOTECH疗法有效抑制实体瘤的生长,并显著提高生存期,为实体瘤的治疗提供了新的策略和希望。

06
该方法在人源化模型中的表现如何?
在该研究中,研究人员首先将患者来源的胰腺导管腺癌(PDAC)肿瘤组织接种到人源化NSG小鼠体内,成功构建了模拟人体肿瘤生物学特性和微环境的人源化PDX模型。接下来,科学人员使用携带前列腺干细胞抗原(PSCA)的293T细胞感染工程化腺病毒(eOA),制备出含有eOA的病毒微囊(MV@eOA)。研究人员利用PSCA特异性CAR-T细胞与MV@eOA中的PSCA相互作用,实现了MV@eOA与CAR-T细胞的物理连接,从而形成了CAR-T-MV@eOA嵌合体。
将制备好的CAR-T-MV@eOA嵌合体通过静脉注射到人源化PDX小鼠体内,观察其对肿瘤生长、生存期、PD-L1表达以及免疫细胞数量等指标的影响,以评估其抗肿瘤作用。为了评估CAR-T-MV@eOA治疗的长期效果,研究人员对人源化PDX小鼠进行了造血干细胞(HSC)重建,并重复以上治疗过程。
通过这一系列精心设计的实验,研究人员证实了CAR-T-MV@eOA嵌合体在人源化PDX模型中表现出显著的抗肿瘤效果。
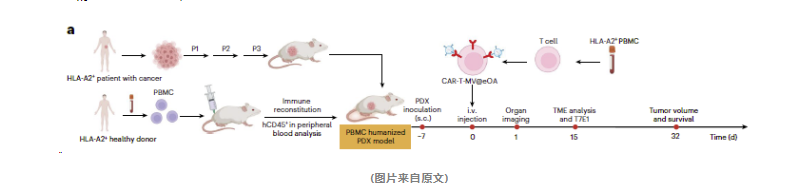
综上所述,ONCOTECH作为一种结合病毒治疗和细胞治疗的技术,展现了良好的临床转化潜力。
翌圣助力产品
在该研究中,研究团队使用了翌圣生物染料法荧光定量qPCR试剂进行病毒拷贝数分析及基因表达分析:

目前翌圣qPCR mix系列的产品已经荣登Nature、 Cell等多个顶级期刊,获得科研大牛们认可!以下仅展示部分助力发表的高分文章:
[1] Li Y, Wang D, Ping X, et al. Local hyperthermia therapy induces browning of white fat and treats obesity. Cell. 2022;185(6):949-966.e19.doi:10.1016/j.cell.2022.02.004.(IF=66.850)
[2] Seki T, Yang Y, Sun X, et al. Brown-fat-mediated tumour suppression by cold-altered global metabolism. Nature. 2022;608(7922):421-428. doi:10.1038/s41586-022-05030-3. (IF=69.504)
[3] Chen P, Wang W, Liu R, et al. Olfactory sensory experience regulates gliomagenesis via neuronal IGF1. Nature. 2022;606(7914):550-556. doi:10.1038/s41586-022-04719-9.(IF=69.504)
[4] Dong W, Zhu Y, Chang H, et al. An SHR-SCR module specifies legume cortical cell fate to enable nodulation. Nature. 2021;589(7843):586-590. doi:10.1038/s41586-020-3016-z.(IF=69.504)
[5] Lu XY, Shi XJ, Hu A, et al. Feeding induces cholesterol biosynthesis via the mTORC1-USP20-HMGCR axis. Nature. 2020;588(7838):479-484. doi:10.1038/s41586-020-2928-y.(IF=69.504)
[6] Bi X, Wang K, Yang L, et al. Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes. Cell. 2021;184(5):1377-1391.e14. doi:10.1016/j.cell.2021.01.046.(IF=66.850)
[7] Liu S, Hua Y, Wang J, et al. RNA polymerase III is required for the repair of DNA double-strand breaks by homologous recombination. Cell. 2021;184(5):1314-1329.e10. doi:10.1016/j.cell.2021.01.048.(IF=66.850)
[8] Liu CX, Li X, Nan F, et al. Structure and Degradation of Circular RNAs Regulate PKR Activation in Innate Immunity. Cell. 2019;177(4):865-880.e21. doi:10.1016/j.cell.2019.03.046.(IF=66.850)
[9] Han X, Wang R, Zhou Y, et al. Mapping the Mouse Cell Atlas by Microwell-Seq. Cell. 2018;172(5):1091-1107.e17. doi:10.1016/j.cell.2018.02.001.(IF=66.850)
[10] Chai Q, Yu S, Zhong Y, et al. A bacterial phospholipid phosphatase inhibits host pyroptosis by hijacking ubiquitin. Science. 2022;378(6616):eabq0132. doi:10.1126/science.abq0132.(IF=63.714)
[11] Yu Q, Liu S, Yu L, et al. RNA demethylation increases the yield and biomass of rice and potato plants in field trials. Nat Biotechnol. 2021;39(12):1581-1588. doi:10.1038/s41587-021-00982-9.(IF=68.164)
[12] Han F, Liu X, Chen C, et al. Hypercholesterolemia risk-associated GPR146 is an orphan G-protein coupled receptor that regulates blood cholesterol levels in humans and mice. Cell Res. 2020;30(4):363-365. doi:10.1038/s41422-020-0303-z.(IF=46.297)
[13] Wang Z, Lu Z, Lin S, et al. Leucine-tRNA-synthase-2-expressing B cells contribute to colorectal cancer immunoevasion. Immunity. 2022;55(6):1067-1081.e8. doi:10.1016/j.immuni.2022.04.017.(IF=43.474)
[14] Bi Q, Wang C, Cheng G, et al. Microglia-derived PDGFB promotes neuronal potassium currents to suppress basal sympathetic tonicity and limit hypertension. Immunity. 2022;55(8):1466-1482.e9. doi:10.1016/j.immuni.2022.06.018.(IF=43.474)
[15] Wang X, Ni L, Wan S, et al. Febrile Temperature Critically Controls the Differentiation and Pathogenicity of T Helper 17 Cells. Immunity. 2020;52(2):328-341.e5. doi:10.1016/j.immuni.2020.01.006.(IF=43.474)
[16] Xiao J, Li W, Zheng X, et al. Targeting 7-Dehydrocholesterol Reductase Integrates Cholesterol Metabolism and IRF3 Activation to Eliminate Infection. Immunity. 2020;52(1):109-122.e6. doi:10.1016/j.immuni.2019.11.015.(IF=43.474)
[17] Zhang X, Zhang C, Qiao M, et al. Depletion of BATF in CAR-T cells enhances antitumor activity by inducing resistance against exhaustion and formation of central memory cells. Cancer Cell. 2022;40(11):1407-1422.e7. doi:10.1016/j.ccell.2022.09.013.(IF=38.585)
[18] Wang XY, Wei Y, Hu B, et al. c-Myc-driven glycolysis polarizes functional regulatory B cells that trigger pathogenic inflammatory responses. Signal Transduct Target Ther. 2022;7(1):105. Published 2022 Apr 18. doi:10.1038/s41392-022-00948-6.(IF=38.104)
[19] Fan H, Hong B, Luo Y, et al. The effect of whey protein on viral infection and replication of SARS-CoV-2 and pangolin coronavirus in vitro. Signal Transduct Target Ther. 2020;5(1):275. Published 2020 Nov 24. doi:10.1038/s41392-020-00408-z.(IF=38.104)
[20] Ren Y, Wang A, Wu D, et al. Dual inhibition of innate immunity and apoptosis by human cytomegalovirus protein UL37x1 enables efficient virus replication. Nat Microbiol. 2022;7(7):1041-1053. doi:10.1038/s41564-022-01136-6.(IF=30.964)
<上下滑动查看更多>
翌圣生物基因研究完整解决方案

产品特别推荐
|
方法 |
分类 |
产品名称 |
货号 |
|
RNA提取 |
同Trizol提取 |
TRIeasy™ Total RNA Extraction Reagent |
10606ES |
|
免氯仿升级版 |
TRIeasy™ Total RNA Extraction Reagent(Tcm Free) |
19202ES |
|
|
动物组织/细胞总RNA提取,避开有毒试剂,最快15 min完成 |
MolPure® Cell/Tissue Total RNA Kit细胞/组织总RNA提取试剂盒 |
19221ES |
|
|
qPCR染料法 |
高特异高荧光值定量预混液(染料法) |
Hieff UNICON® Advanced qPCR SYBR Master Mix |
11185ES |
|
高灵敏通用型定量预混液(染料法) |
Hieff UNICON® Universal Blue qPCR SYBR Master Mix |
11184ES |
|
|
超高性价比定量预混液 (染料法),已发文章累计IF达到5000+ |
Hieff® qPCR SYBR Green Master Mix (No Rox) |
11201ES |
|
|
Hieff® qPCR SYBR Green Master Mix (Low Rox) |
11202ES |
||
|
Hieff® qPCR SYBR Green Master Mix (High Rox) |
11203ES |
||
|
反转录试剂 |
5 min一步gDNA去除&反转录预混液(下游应用qPCR) |
Hifair® AdvanceFast One-step RT-gDNA Digestion SuperMix for qPCR |
11151ES |
|
5 min快速反转,最长可满足14 kb cDNA合成,含gDNA去除(下游应用PCR/qPCR) |
Hifair® AdvanceFast 1st Strand cDNA Synthesis Kit |
11149/11150ES |
|
|
高质量第一链cDNA合成预混液,含gDNA去除(下游应用qPCR) |
Hifair® III 1st Strand cDNA Synthesis SuperMix for qPCR (gDNA digester plus) |
11141ES |